
9870 St Vincent Place, Glasgow, DC 45 Fr 45.

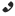
Вид: Elaphe situla = Леопардовый полоз

- Класс: Reptilia = Пресмыкающиеся
- Подкласс: Lepidosauria = Лепидозавры, чешуйчатые ящеры
- Отряд: Squamata Oppel = Чешуйчатые
- Подотряд: Serpentes {Ophidia} Linnaeus, 1758 = Змеи
- Семейство: Colubridae Cope = Ужеобразные змеи, ужовые
- Род: Elaphe Wagler , 1833 = Полозы (четырехполосые)
- или: Elaphe Fitzinger, 1833 = Лазающие полозы, крысиные змеи
- Вид: Elaphe situla (Linnaeus, 1758) = Леопардовый полоз
Леопардовый полоз в Крыму.
РАСПРОСТРАНЕНИЕ И ЭКОЛОГО - МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ЛЕОПАРДОВОГО ПОЛОЗА В КРЫМУ
О. В. Кукушкин, А. Н. Цвелых

Исследованы распространение, численность, динамика сезонной и суточной активности, отношение к температуре, другие особенности биологии, а также морфология леопардового полоза в Крыму. Вид повсеместно распространен на южном макросклоне, в западной и восточной частях северного макросклона Крымских гор до высоты 680 м над ур. м., а также на Гераклейском п-ве. Отмечено постоянное обитание в населенных пунктах, включая крупные города (г. Севастополь). Обычно встречаются единичные особи, но в оптимальных биотопах численность может достигать 2.3-8.9 экз/га. Количество самцов превышает количество самок в 1.7 раза. Активен с марта по ноябрь. Во время оттепелей встречается и зимой. Максимальная активность наблюдается в период размножения. Ведет ночной и дневной образ жизни. Ночная активноcть максимальна в августе. Активен при температуре воздуха 16–29°С, субстрата – 16–35°С. Спаривание происходит в апреле-мае, кладка - в июне-июле. Линька - с марта по ноябрь. В питании отмечены мелкие млекопитающие, птицы и ящерицы. Половой диморфизм проявляется в общих размерах, количестве брюшных и подхвостовых щитков, соотношении размеров тела и хвоста. Доля полосатой формы максимальна в центре ареала. На периферии ареала резко преобладают особи пятнистой морфы.
Леопардовый полоз - чрезвычайно редкий, находящийся на грани исчезновения вид (Ананьева и др., 1998), включенный в Красные книги Международного Союза Охраны Природы и Украины, является одним из наименее изученных представителей герпетофауны Восточного Средиземноморья. Имеющиеся на сегодняшний день сведения о леопардовом полозе в Крыму крайне скудны, неполны и нередко противоречивы.
МАТЕРИАЛ И МЕТОДИКА
Исследования проводились в период 1989-2003 гг. Питание змей изучали исключительно бескровными методами: при анализе отрыгнутой добычи, при непосредственных наблюдениях за змеями в природе, а также при вскрытии змей, найденных погибшими. Пол у погибших змей определяли при вскрытии, у живых - зондированием. С разной степенью полноты использованы данные о 194 особях.
Распространение:Традиционно принято считать (Щербак, 1966, 1988), что в Крыму леопардовый полоз распространен почти исключительно на южном макросклоне Главной гряды гор и проникает в предгорья лишь на Гераклейском п-ве, в ближайших окрестностях Севастополя. Результаты наших исследований подтвердили и дополнили существующие представления о характере распространения этого вида в центральной части крымского ареала. Однако в западной части полоз оказался намного шире распространен, чем это было известно ранее. Здесь он населяет как южный, так и северный макросклоны Главной гряды Крымских гор, низко- и среднегорные местности в пределах второй гряды; наиболее северная находка - гора Чуфут-Кале под Бахчисараем. Леопардовый полоз населяет также практически весь Гераклейский п-в. На восток вид прослежен до г. Феодосии (западные окраины города и хребет Тепе-Оба) (О. Г. Розенберг и М. М. Бескаравайный, личные сообщения). Северная граница распространения полоза в восточной части ареала проходит через г. Старый Крым (О. Г. Розенберг, личное сообщение).
В западном предгорье и на северо-востоке Байдарской долины вид прослежен до высоты 520 м над ур. м., на крайнем западе Главной гряды - до 680 м над ур. м. (хребет Кокиябель и гора Ильяс-Кая). Приблизительно до такой же высоты (600 м над ур. м.) вид был отмечен и в восточной части ареала (Щербак, 1966).
Биотопы:В Крыму E. situla населяет скалисто-каменистые сухие высокоможжевеловые, пушистодубово-фисташковые редколесья и сухие можжевелово-сосновые сугрудки, томилляры, мергелистые и шиферные склоны в различного рода шибляках, мелкоствольные грабинниковые и кизиловые дубняки, окраины дубово- и грабово-буковых (иногда с участием сосны Палласа или тиса) лесов, прирусловые леса из ясеня, ольхи, ивы, рудеральные местообитания, иногда окраины виноградников и плантации розы. У верхней границы шиблякового пояса (500-600 м над ур. м.) обнаружен в низкорослых редколесьях из ясеня и лохолистной груши. Предпочитает каменистые биотопы с плотными зарослями кустарника (иглица понтийская, жасмин кустарниковый, ладанник крымский, боярышники и др.). Встречается даже на сильно деградировавших от воздействия антропогенного фактора участках побережья близ часто посещаемых пляжей. Обычно встречается на небольшом (до 300 м) удалении от воды (реки, ручьи, родники). Как правило, придерживается склонов южной, юго-западной, юго-восточной и западной экспозиций, но встречен и на склонах северной и северо-восточной экспозиций. Причем обитание на северных склонах отмечено и в климатически наиболее суровых частях ареала: у крайних северных границ обитания вида в юго-западном Крыму, а также на Карадаге и расположенном к западу от него Эчкидаге (Щербак, 1989; М. М. Бескаравайный, личное сообщение).
Убежищами полозам служат трещины скал, полости между камней, кладки стен из бута, старые или свежевырытые норы грызунов. Причем норы могут располагаться как на травянистых, так и на совершенно лишенных растительности эродированных участках склона.
Полозы нередко встречались в черте населенных пунктов (Щербак, 1966, 1989; Бескаравайный, 1987). Тенденция к синантропизации проявляется в том, что вид охотно заселяет различного рода развалины и даже жилые строения. Полозов неоднократно отмечали в крупных городах: Севастополе, Ялте, Алуште, Судаке. По данным С. А. Шарыгина (личное сообщение), полоз встречается в подпорных каменных стенах даже в центре Ялты. Особо следует сказать об обитании этого вида в г. Севастополе, тем более что и ранее его здесь находили в городских постройках (Щербак,1966). Полоз нередко встречается в городских кварталах одноэтажной послевоенной застройки, особенно если по соседству есть плотные заросли колючих кустарников (дереза, ежевика). Осенью (в октябре - ноябре), в поисках зимовочных убежищ, полозы нередко проникают в жилые дома, сараи (см. также Щербак, 1966), погреба, и даже в систему городской канализации. Благодаря особенностям городской застройки, где жилые массивы разделяются балками, оврагами, склонами, поросшими кустарниковой или древесной растительностью, полозы могут постоянно обитать в центре города.
Антропогенный фактор является наиболее частой причиной гибели полозов. Полозы регулярно гибнут от прямого преследования человеком (отмечено также несколько случаев гибели полозов под колесами автомобилей). На территориях, испытывающих мощный антропогенный пресс, нередко встречаются травмированные особи - обычно у них отсутствует часть хвоста. Следует заметить, что влияние антропогенного фактора не является решающим для выживания змей этого вида - очень крупные особи (с длиной тела более 850 мм) неоднократно встречались как в центре Севастополя, так и на сильно вытоптанных приморских террасах вблизи Балаклавы.
Численность:Широко распространено мнение о крайне низкой и неуклонно снижающейся численности леопардового полоза (Щербак, 1977, 1988, 1994). При этом обычно игнорируется иное, базирующееся на многолетних стационарных наблюдениях в центральной части ареала мнение, согласно которому численность полоза в Крыму далеко не столь катастрофически низка (Шарыгин, 1983) и достаточно стабильна (Шарыгин, 1985).
Как правило, встречается спорадично, единичными особями. Максимальная численность - 4 экз. на 1.5 км маршрута (2.67 экз/км или 8.9 экз/га) - зарегистрирована 2.V 1997 г. под пологом высокоствольных дубово-грабинниковых шибляков на скальных карнизах севернее мыса Айя. В Чернореченском каньоне (1-2.VI 1997, 13-14.VI 1999), в ущельях рек Демерджи, Алак и Сотера (10-11.VI 1997), на мысе Мартьян (13.V 1999), на горе Аюдаг (28.X 2000) в оптимальных биотопах, на трехкилометровых маршрутах учитывали по 2 экз. (0.7 экз/км или 2.3 экз/га). В руинах заброшенного пос. Узен-Баш (Хворостянка) к востоку от с. Родное в третьей декаде апреля 1998 г. отмечена плотность 5 экз/га, а в руинах близ ст. Верхнесадовая в конце сентября 1990 г. (А. А. Алексеев, личное сообщение) - 5 взрослых змей на площади 100-120 кв. м. Возможно, в последнем случае наблюдалось предзимовочное скопление.
Сезонный цикл активности:Считается, что в Крыму полозы выходят из зимней спячки во второй половине апреля (Щербак, 1966; Ананьева и др., 1998). По нашим данным наиболее ранние находки змей этого вида на поверхности приходятся на конец февраля - первую декаду марта: 24.II и 1.III 1998 (2.5 км к юго-востоку от Балаклавы); 7.III 1999, (2.5 км к востоку от с. Родное). Возможно, в отдельные годы сроки появления полозов в горах могут значительно смещаться, например первая встреча активных полозов в последнем из пунктов в 1998 г. относится только к 13.IV 1998 (спустя 2 - 3 дня после таяния снега). Окончательно уходит на зимовку не ранее середины ноября: по всему ареалу активные полозы достаточно часто встречаются во второй половине октября - первой половине ноября. Последние даты встреч - 19.XI 1982, 20.XI 1985 (Карадаг) (Щербак, 1989; М. М. Бескаравайный, личное сообщение), 21.XI 2002 (в черте г. Севатополя, наши данные) и 24.XI 1993 (между Балаклавой и мысом Айя, наши данные). Полозы изредка появляются на поверхности и в зимний период - во время оттепелей. Зимой они отмечались 12.XII 1999 и 31.XII 2000 (окрестности Севастополя); 16.I 1998 (мыс Фиолент); 5.II 1999, (окрестности г. Алушта, В. В. Соловьев, личное сообщение).
После пробуждения активность полозов постепенно нарастает (рис. 2) и достигает наиболее высокого уровня в мае, что совпадает с периодом размножения. Далее, в летние месяцы активность закономерно снижается достигая минимума в наиболее жаркий период - в июле-августе. С наступлением осени полозы вновь начинают встречаться несколько чаще. Очевидно, уменьшение общего количества встреч полозов в самые жаркие месяцы частично может объясняться переходом животных преимущественно к ночному типу активности (см. ниже).
Суточная активность:В зависимости от сезона для полоза характерны как дневной, так и ночной типы активности. С момента пробуждения и до конца апреля регистрируется исключительно дневная активность. Затем, с увеличением среднесуточной температуры и уменьшением влажности воздуха постепенно повышается уровень сумеречно-ночной активности: отношение количества особей встреченных в светлое время суток и встреченных вечером, изменяется от 3.67:1 в мае до 0.67:1 в августе (рис. 3). Круглосуточные наблюдения, проведенные в середине лета (13-14.VII 1998) за индивидуально опознаваемой (по деталям окраски) особью (взрослый самец, окрестности с. Родное), позволили выявить 2 фазы активности: дневную - между 9 и 14 ч (при этом змея в основном грелась, не удаляясь от убежища далее чем на 1.5 м) и ночную - после 21 ч (полоз покинул убежище в нише стены в 21 ч 15 мин). Продолжительность ночной активности и радиус ночных передвижений проследить не удалось, однако в 9 ч утра следующего дня греющийся полоз уже был обнаружен в освещенном убежище неподалеку от входа.
Ночные дожди стимулируют повышение дневной активности. В целом дневной активности наиболее благоприятствует влажная нежаркая облачная погода. Во второй половине лета, при установлении сухой и жаркой погоды полозы переходят преимущественно к сумеречно-ночному образу жизни, осенью они наиболее активны днем. В сентябре-ноябре в дневное время полозы встречались в 5 раз чаще, чем в сумеречно ночные часы (n=24). Последние находки змей в сумерках приходятся на вторую декаду октября (15.X 1998, Севастополь, Херсонес) - первую декаду ноября (4.XI 1998, окрестности Балаклавы). В целом в течение года в сумерках и ночью полозы встречались в 3.83 раза реже, чем в дневные часы.
Отношение к температуре:На протяжении активного периода змеи отмечались на поверхности днем между 9 и 15 ч при температуре воздуха 16.0-28.6°С, субстрата - 16.2 (обычно выше 19.5°С) –35.2°С и в темное время суток между 17 ч (в конце осени) – 20 ч 30 мин (летом) и полуночью при температуре воздуха и субстрата соответственно 16(обычно выше 19.5°С) - 25°С и 19 (обычно выше 21°С) – 25.5°С.
В марте-мае, при температуре воздуха около 20°С полозы обычно начинают проявлять активность только после того, как температура в убежищах поднимается выше 15°С. При температуре воздуха 14-15°С, субстрата - 16-18°С и ясной погоде змеи укрытий не покидали. Слабую активность полозы могут проявлять и при более низких температурах: в апреле 1999 г. в Мекензиевском лесничестве (окрестности Севастополя) в первой половине дня (переменная облачность) при температуре воздуха всего 7.5°С отмечен наполовину вылезший из убежища малоактивный полоз (Ю. В. Кармышев, личное сообщение).
В целом в течение года подавляющее большинство активных полозов встречено в диапазоне температур воздуха и субстрата 19-26°С. Летом, в сумерках и ночью, змеи встречаются преимущественно при температуре воздуха и субстрата 24.5-26°С. Во время уже упоминавшихся круглосуточных наблюдений 13-14.VII 1998 отмечено, что вечером полоз покинул убежище в нише стены при температуре воздуха и субстрата 21.9°С. При этом температура в убежище составляла 24.8°С.
Осенью, во второй декаде октября - первой декаде ноября в сумерках змеи встречались при температуре воздуха 16-17°С, субстрата – 19-21°С. В полдень 29.X 2000 на горе Аюдаг греющаяся на выходе из убежища особь найдена при температуре воздуха и субстрата 16.2°С.
Взрослая самка, находившаяся несколько суток в террариуме (с 1.XI 2001) при температурах, близких к естественным (суточный максимум 12-14°С, минимум 5-10°С), проводила все светлое время суток в убежище, покидая его только с наступлением темноты (около 17 ч) при температуре 9.2-11.5°С, при этом она была малоподвижна. При экспонировании террариума на солнце полоз проявлял слабую активность между 12 и 13 ч при температуре 18.5-21°С, но не покинул убежище в темное время суток при температуре 9.1-10.3°С.
Размножение:Неоднократно отмечалось (Щербак, 1966, 1977, 1994), что количество самцов в популяции леопардового полоза в Крыму значительно превышает (по некоторым данным более чем в 4 раза - Щербак,1966) количество самок. Наши данные также подтверждают более редкие встречи самок по сравнению с самцами, однако диспропорция в отношении взрослых самцов и самок в нашей выборке оказалась менее явной: 1.71:1 (n = 38). Можно было бы допустить, что на самом деле эта диспропорция объясняется большей подвижностью взрослых самцов и (или) большей скрытностью самок у этого вида. Однако сходные данные получены нами и для молодых неполовозрелых (длиной до 400 мм) леопардовых полозов: 1.75:1 (n=11). Таким образом, общее отношение полов в нашей выборке 1.72:1 (63% самцов, 37% самок, n = 49).
Судя по срокам откладки яиц в неволе (см. ниже), спаривание в годы с ранней весной может начаться во второй декаде апреля и, по-видимому, продолжается до конца мая (максимальные размеры семенников отмечены у самца, добытого в первой половине мая (Щербак,1966)). Только в этот период регулярно встречались пары полозов. Иногда удавалось наблюдать следование самца за самкой, однако копуляция не была отмечена ни разу. Крайние даты встреч таких пар 13.IV 1998 - 11.V 1991.
Кладки леопардового полоза в естественных условиях в Крыму до сих пор не найдены. Однако сведения о более мелких размерах яиц крымских E. situla (Ананьева и др., 1998), вероятно, не соответствуют действительности, так как использованы данные измерений яиц, извлеченных из яйцеводов змей (Щербак, 1966), к тому же хранившихся в фиксирующем растворе более 50 лет. Размеры таких яиц действительно невелики: 35-37.7x9.5-12 мм (Щербак, 1966). Измерения яиц из кладок показало, что они значительно крупнее. По данным Шарыгина (1986), самка, отловленная 14.VII 1982 близ пос. Ливадия, 22 июля отложила 3 белых яйца размером 50-60x15-20 (в среднем 55.3x17.3) мм. По нашим данным самка, отловленная 22.IV 1996 близ с. Родное, 1.VII отложила 3 белых яйца имевших размеры 46--54x17-22 (в среднем 50.7x19.0) мм. Особь, отловленная 2.V 1997 близ мыса Айя, 15.VII отложила очень крупное (78.8x17.3 мм) неоплодотворенное яйцо с несформированной скорлуповой оболочкой. Приведенные сроки откладки яиц (есть также сообщение О. Б. Спивакова о факте откладки двух яиц в июле самкой, содержавшейся непродолжительное время в террариуме карадагского экзотариума) в общем согласуются со сроками, предполагаемыми на основании анализа состояния гонад крымских леопардовых полозов (Щербак, 1966): конец июня - начало июля.
Во всех случаях откладка яиц происходила ночью (в двух последних - около полуночи). В обоих случаях нормальной кладки поведение самок было сходным: они закрывали кладку (яйца в кладке ориентированы вертикально) кольцами своего тела, держась поверх яиц, и вели себя крайне агрессивно. При этом температура кладки существенно превышала температуру окружающей среды (Шарыгин, 1986). Таким образом, есть основания предполагать, что самки леопардового полоза охраняют и, возможно, согревают кладку.
Продолжительность инкубации в естественных условиях остается неизвестной. Выход молодых полозов из яиц приходится, по всей видимости, на вторую половину августа - сентябрь, так как первые встречи сеголеток приходятся на август.
Питание:Полозы начинают питаться сразу после пробуждения: в экскрементах змей, собранных в конце февраля - начале марта 1998 г., обнаружена шерсть полевок. Нами исследованы 14 проб питания змей разного пола и возраста (L.=395-850 мм). Учтены также опубликованные (Щербак, 1966, 1989) результаты анализа еще 4 пищевых проб, где объекты питания удалось определить с достаточной точностью (до рода). При этом в отдельных случаях видовую принадлежность млекопитающих корректировали с учетом современных данных о видовом составе териофауны в пределах ареала леопардового полоза в Крыму. Например, здесь зарегистрировано только по одному представителю родов Sorex и Microtus.
В рационе взрослых полозов превалируют (отмеченные в питании у 86 % особей (n=15)) мелкие млекопитающие (табл. 2). Питание птицами отмечено у взрослых змей в трех случаях. В одном из них в желудке полоза обнаружена взрослая птица (зарянка), а в двух остальных - наблюдалось поедание птенцов (коноплянка, зарянка). Причем в последнем случае у гнезда птиц наблюдалась пара полозов, но птенцов (всего в выводке было 5 птиц в возрасте около 7 дней) заглатывала только самка. Впоследствии, кроме двух птенцов, эта особь отрыгнула еще и съеденную накануне молодую желтогорлую мышь. Молодое животное этого вида отмечено также в питании E. situla в Карадаге (Щербак, 1989). Наиболее крупный объект, зарегистрированный в питании полозов, - взрослая желтогорлая мышь, проглоченная змеей длиной 715 мм.
Рептилии - некрупные скальные ящерицы (Darevskia lindholmi) и взрослая крымская ящерица (Podarcis taurica) - отмечены в питании только у молодых полозов. Сведения о питании сеголеток леопардового полоза голыми слизнями и дождевыми червями (С. А. Шарыгин, личное сообщение) не потверждаются нашими наблюдениями. По нашим данным, а также по данным Н.Н. Барвинского (личное сообщение), в условиях террариума молодые полозы отказываются от беспозвоночных и питаются исключительно новорожденными мышатами или ящерицами.
Линька:Литературные данные о линьке E. situla в Крыму отсутствуют. Самка из окрестностей с. Родное, отловленная в конце апреля 1996 г., в неволе полиняла 6.V. Взрослых змей на поздних стадиях линьки (с помутневшими брюшными и глазными щитками) встречали 1.III 1998 (окрестности Балаклавы, самка), 13.IV и 13.VII 1998 (гора Джилен, самец), 6.VI 1998 (гора Парагильмен), 12.VII 2001 (Карадагский заповедник, самка), 9.X 2001 (гора Ильяс-Кая), 19.V 2002 (Севастополь, самец), 29.V 2002 (Севастополь, самец); линяющих - 28.V 1998 (каньон р. Уркуста), 1.VI 1997 (каньон р. Черная), 19.V 2002 (Севастополь, самец); на начальных этапах линьки (с помутневшими брюшными щитками) - 19.V 2002 (Севастополь, самец). Змеи с очень ярким контрастным узором, по видимому, недавно перелинявшие, встречены: 7.III 1999 и 16.IV 1998 (окрестности с. Родное, самки), 4.IV 1999 (мыс Фиолент, самец); 2.VII 1998 (гора Ильяс-Кая, самец), 13.V 1999 (мыс Мартьян, самец), 28.X 2000 (гора Аюдаг), 1. XI 2001 (мыс Фиолент, самка), 14.V 2002 (Севастополь, самка) и 23.VIII 2002 (Варнутская долина, самец).
Поведение:Во время охоты полозы активно обследуют различные убежища, включая довольно труднодоступные: змей, обследующих трещины, в сумерках и ночью наблюдали на почти отвесных скалах на высоте до 1,8 м от подножья.
Обороняясь, E. situla, подобно некоторым другим видам лазающих полозов (Банников и др., 1977), способен вибрировать кончиком хвоста. Эта поведенческая реакция наблюдалась у четырех самцов в апреле-мае.
Леопардовый полоз не избегает соседства с другими рептилиями: нередко встречается в одних биотопах с желтобрюхим (Coluber caspius) и палласовым (Elaphe sauromates) полозами, а также желтопузиком (Pseudopus apodus). Во время круглосуточных наблюдений в июле в нише небольшого (площадью около 1,5 кв. м) фрагмента разрушающейся стены, укрепляющей склон, где обитали леопардовые полозы, также удавалось попеременно наблюдать крупных палласова и желтобрюхого полозов (днем) и желтопузика (ночью).
Морфологическая характеристика:Максимальные размеры полозов нашей выборки (L.=908 мм, L. cd.=215 мм) существенно превышают приводившиеся ранее максимальные для Крыма размеры (Щербак, 1966): 780 и 194 мм, соответственно. Однако размеры крымских леопардовых полозов все же значительно меньше, чем приводимые для вида в целом: L.=1160 мм, L.cd.=350 мм (Ананьева и др., 1998). Диапазон изменчивости (табл.3) относительной длины хвоста, количества брюшных (у самцов) и подхвостовых чешуй (у самок) значительно превышает диапазон изменчивости этих параметров в выборке крымских леопардовых полозов, исследованной Щербаком (1966). Диапазон изменчивости каждого из остальных параметров в общем соответствует диапазону изменчивости параметров этой выборки. Из 38 полозов нашей выборки 83,3 % имело 27 чешуй в ряду вокруг середины туловища (Sq.), у 14,3 % их было 25 и только одна особь (2,4 %) имела 23 чешуйки в ряду.
Как и у многих других видов рода Elaphe, половой диморфизм у леопардового полоза проявляется в общих размерах, соотношении количества брюшных щитков и пар подхвостовых щитков и в соотношении размеров тела и хвоста (табл.3). Взрослые самки в общем несколько крупнее самцов: у более чем половины (54%, n = 13) осмотренных самок длина тела превышала 800 мм, в то время как среди самцов таких особей было только несколько больше трети (38%; n = 16). Достоверные половые различия отмечены в количестве брюшных и подхвостовых щитков, в их соотношении и в длине хвоста по отношению к длине тела (табл.3).
Известно, что в Крыму встречаются полозы двух вариаций: полосатой - var. situla и пятнистой - var. leopardina. Изредка встречаются особи с промежуточным типом рисунка, сочетающим элементы обеих форм (Щербак, 1966): за время исследований нами отмечены только пять таких особей, что составляет 3.6 % от всей изученной выборки. В центральной части ареала - на Южном берегу Крыма, в районе Ялты - особи полосатой морфы, по-видимому, преобладают (Шарыгин, 1983). По данным Щербака (1966) касающихся, судя по всему, также преимущественно полозов южного макросклона, меньше половины исследованных змей принадлежали к пятнистой морфе. Анализ встречаемости морф в центральной части ареала - на Южном берегу Крыма (приморский склон от района Балаклавы и мыса Айя на западе до района г. Судак на востоке) показывает, что здесь действительно преобладают особи полосатой морфы (табл. 4). Совершенно иным оказалось соотношение морф в других частях ареала. У восточной границы распространения вида (горные и предгорные районы восточнее г. Судак) особи пятнистой морфы встречаются в 5.5 раза чаще, чем полосатой. То же наблюдается и в западной части ареала: на северном макросклоне Главной гряды и в предгорьях особи вариации leopardina встречаются в 4.08 раза чаще, чем situla. Причем если анализировать отдельно окраску животных из районов, непосредственно прилегающих к северному макросклону Главной Гряды (так называемое Байдарское низкогорье, включающее Байдарскую и Варнутскую долины, а также долину Хайту и Чернореченский каньон), и районов, расположенных еще западнее (предгорья, включая Гераклейский п-в), то можно увидеть (табл. 4, рис.4), что доля пятнистой морфы еще более возрастает у наиболее западных популяций (соответственно, 2.20 и 5.25).
Таким образом, доля полосатой формы леопардового полоза в Крыму минимальна на периферии ареала и достигает максимума в центральной части ареала (Южный берег Крыма), которая отличается наиболее мягким климатом и, по-видимому, является наиболее древней его частью.
Заключение
Трудно предположить, что ареал леопардового полоза в Крыму стремительно расширился за последние полвека. По-видимому, во многих районах этот скрытный вид просто не отмечался исследователями ранее, чему возможно способствовала его низкая численность и (или) естественная мозаичность ареала близ северных границ распространения в Крыму. Например, первая находка леопардового полоза на Карадаге – районе, хорошо изученном зоологами, позволившая существенно сдвинуть восточную границу ареала этого вида в Крыму, произошла только в 1980 г. (Щербак, 1989).
Приведенные данные по распространению и биологии леопардового полоза свидетельствуют о большей, чем предполагалось, экологической пластичности этого реликтового стенобионтного вида. Так, Байдарская котловина, где леопардовый полоз распространен достаточно повсеместно, в сравнении с Южным берегом Крыма и Гераклейским п-вом, отличается заметно более прохладным климатом. Предполагается (Ена, 1990), что сходные условия могли существовать в неоплейстоцене в приморских рефугиумах реликтовой средиземноморской растительности на Южном берегу Крыма. Можно предположить, что в тот период ареал полоза был минимальным и впоследствии расширился с изменением климатических условий в регионе. Не исключено, что в связи с общим потеплением расширение ареала леопардового полоза происходит и в настоящее время, поскольку современные климатические условия в принципе благоприятствуют увеличению численности и распространению этого термофильного вида.
Широко распространенное мнение о чрезвычайной редкости леопардового полоза в Крыму (Щербак, 1966,1988, 1994; Ананьева и др., 1998) сильно преувеличено. Как справедливо отмечает Шарыгин (1983, 1985, 1998, 1998 а), исчезновение этому скрытному виду не угрожает. Помимо заповедных территорий естественными резерватами вида в настоящее время являются относительно слабо трансформированные хозяйственной деятельностью человека западные предгорья к югу от р. Бельбек и, в особенности, труднодоступные и малопосещаемые людьми скалистые урочища крайнего запада Главной гряды.
СПИСОК ЛИТЕРАТУРЫ
Ананьева Н.Б., Боркин Л.Я., Даревский И.С, Орлов Н.Л.,1998. Земноводные и пресмыкающиеся. М.: ABF. 576 с. Банников А.Г., Ищенко В.Г., Рустамов Э.К., Щербак Н.Н., 1977. Земноводные и пресмыкающиеся фауны СССР. М.: Просвещение. 415 с. Бескаравайный М.М., 1987. Земноводные и пресмыкающиеся // Карадагский государственный заповедник АН УССР. Летопись природы. Т. 4. Кн. 2. С. 63. Даревский И. С., 1987. Методы изучения рептилий в заповедниках // Амфибии и рептилии заповедных территорий. М. С. 25-35. Даревский И.С., Щербак Н.Н., Татаринов К.Н., Ищенко В.Г. и др., 1989. Руководство по изучению земноводных и пресмыкающихся. Киев: Наукова думка. 172 с. Ена А.В., 1990. Реликтовый земляничник // Природа. № 12. С. 42-48. Кармышев Ю.В., 2001. Новые данные о распространении леопардового полоза (Elaphe situla) в Крыму // Вестн. зоол. №1. С. 52. Шарыгин С.А., 1983. К изучению редких видов герпетофауны Крыма // Охрана живой природы. Тез. докл. Всес. конф. молодых ученых (ноябрь 1983 г.). М. С. 212 - 214. - 1985. Охрана герпетофауны в заповедниках Крыма // Теоретические основы заповедного дела. Тез. докл. Всес. совещ. (Львов, 18-19 декабря 1985 г.). М. С. 304-307. - 1986. Кладка леопардового полоза в неволе // Первое Всес. совещ. по проблемам зоокультуры. Тез. докладов. Ч. 2. М. С. 163-165 – 1998. Сохранение герпетофауны на заповедных территориях Крыма // Состояние природных комплексов Крымского природного заповедника и других заповедных территорий Украины, их изучение и охрана. Алушта. С. 94-95. - 1998 а. Рептилии и амфибии в заповедниках Крыма // Роль охоронюваних природних территорий у збереженнi бiорiзноманіття. Канiв. С. 255-256. Щербак Н.Н., 1966. Земноводные и пресмыкающиеся Крыма. Киев.: Наукова думка. 240 с. - 1977. Страницы "Красной Книги" Леопардовый полоз // Природа. №12. С. 103-104 - 1988. Пресмыкающиеся // Редкие и исчезающие растения и животные Украины. Киев.: Наукова думка. С. 160-165. - 1989. Земноводные и пресмыкающиеся // Флора и фауна заповедников СССР. Фауна Карадагского заповедника. М. С. 33-37. - 1994. Полоз леопардовий // Червона книга України.Тваринний світ. Київ: Українська енциклопедія. С. 297. Кукушкин О. В., Цвелых А. Н. Распространение и эколого-морфологические особенности леопардового полоза, Elaphe situla (Serpentes, Colubridae), в Крыму // Зоологический журнал. – 2004. – 83, N 4. – С. 439 – 448.