
9870 St Vincent Place, Glasgow, DC 45 Fr 45.

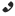
Главная сайта "Мир Животных" / Ваш аквариум / Аквариумные растения / Каталог аквариумных рыб / Биология рыб / Все о рыбалке /
Болезни рыб / Самые интересные рыбы /
Целакант: Поведение

Читать: Целакант ;
Целакант: Поведінка
При перечікуванні денного часу целаканти збираються досить великими групами: одного разу в підводній печері було нараховано 19 дорослих риб, що повільно рухались за допомогою парних плавців, при цьому ніколи не торкаючись одна одної. Особини, що їх ідентифікували за розташуванням світлих плям, на протязі багатьох місяців траплялись в одних і тих самих печерах; але знаходились і такі, що міняли печери кожного дня. Вночі всі риби поодинці переміщуються ближче до поверхні.

Вже після перших спостережень в 1987 році при зануренні батискафу GEO, біолог Ганс Фріке відзначив, що вночі всі целаканти дозволяють переносити себе висхідними та низхідними течіями води, а також горизонтальними течіями. Парні плавці стабілізують дрейфуючу рибу таким чином, що вона заздалегідь обминає будь-які перешкоди. Також Фріке повідомив (і в подальшому це спостереження підтвердилося), що всі риби час від часу перевертаються вертикально головою до низу, і лишаються в цьому положенні до двох хвилин. Сенс такої риси поведінки досі невідомий.
При плаванні целакант повільно рухає парними лопасними грудними та черевними плавцями в протиставному порядку, тобто одночасно лівим грудним та правим черевним, а потім одночасно правим грудним та лівим черевним. Такі рухи характерні також для легеневої риби та деякої (дуже малої) кількості інших видів риб, що ведуть придонний спосіб життя, але не для нектонних риб. Окрім того, цей спосіб руху кінцівок є основним для сухопутних хребетних.
Непарні лопасні другий спинний та анальний плавці коливаються синхронно з боку в бік, і є основним засобом відносно швидкого пересування в передньому напрямі. Це пояснює їхню однакову форму та зеркальне розташування. Нелопасний (променевий) перший спинний плавець звичайно складений вздовж спини, але риба розправляє його при відчутті небезпеки; також цей плавець може використовуватись як вітрило при дрейфуванні з течією.
Великий хвостовий плавець (точніше, злиті третій спинний, хвостовий та другий анальний плавці) перебуває випрямленим та нерухомим на протязі дрейфування або повільного плавання, що характерне для всіх слабо електричних риб: це дає змогу проводити інтерпретацію збуджень навколишнього електричн ого поля. Але у випадку небезпеки хвостовий плавець використовується для швидкого ривку вперед.
ого поля. Але у випадку небезпеки хвостовий плавець використовується для швидкого ривку вперед.
Маленький епікаудальний лопастевидний плавець згинається з одного боку до іншого при русі риби, а також при «стоянні на голові», і може брати участь в електрорецепції разом з ростральним та ретікулярними органами. Команда батискафу GEO змогла викликати у целакантів «стояння на голові» шляхом пропускання слабких електричних токів між електродами, що їх тримав зовнішній маніпулятор.
Описна вище координація руху плавців, вірогідно, еволюційно розвинулась для стабілізації положення громіздкого тіла риби, але у їхніх вимерлих попередників могла сприяти пересуванню по суходолу. При спостереженнях коморського целаканту в природі його парні лопастевидні плавці при контакті з субстратом не використовувались. Тобто, вірогідно, нині живучі целаканти ніколи не «ходять».
Харчування
Індонезійський целакант. На знімку видно коричневе, на відміну від живого коморського целаканта, забарвлення.
Коморський целакант пристосований до нічного придонного харчування з повільним рухом. Відповідні дослідження визначили в якості об'єктів харчування бентичні та навколобентичні організми, такі як анчоуси-світлячки (Myctophidae), глибоководні риби-кардінали (Apogonidae), каракатиці та інші головоногі, люциани, і навіть головату акулу (Cephaloscyllium). Більшість з цих об'єктів харчування переховується у підводних печерах.
Анатомія черепа целакантів (внутрішньочерепний суглоб) обумовлює їхній спосіб добування їжі шляхом всмоктування разом з водою при різкому відкритті рота; таким чином риби «висмоктують» здобич із пустот та розселин в скелях.
Розмноження
До 1975 року коморський целакант вважався яйцекладучим (овіпорозним — див. статтю «Розмноження риб»), позаяк в організмі 163-сантиметрової самки, виловленої біля острову Анжуан в 1972 році, було знайдено 19 яєць, що формою та розміром нагадували апельсин. Але пізніше інша самка, довжиною 160 сантиметрів, що була виловлена біля Анжуану в 1962 році, і перебувала в експозиції Американського Музею Природної Історії (American Museum of Natural History, AMNH) була вскрита в 1975 році. Співробітники музею провели це вскриття для взяття проб тканин внутрішніх органів, і виявили при цьому в яйцеводах самиці п'ять добре розвинутих ембріонів довжиною 30-33 сантиметри, кожний з великим жовтковим міхуром. Це відкриття свідчить, що целаканти є живорідними, тобто розмножуються вівіпорозом (див статтю «Розмноження риб»).
Пізніше дослідники Джон Вурмс та Джим Антц докладно вивчили ембріонів та яйцеводи, і показали, що сильно васкуляризована поверхня жовткового міхура знаходиться в дуже тісному контакті з так само сильно васкуляризованою поверхнею яйцеводу, формуючи плацентоподібну структуру. Таким чином, виглядає вірогідним, що на додаток до жовтка з яйця ембріони також харчуються і завдяки дифузії поживних речовин з крові матері.
Третій можливий шлях живлення був досліджений після вилову та вскриття ще кількох самок коморського целаканту. Одна зних, довжиною 168 см, мала 59 яєць розміром з куряче, інша мала 65 яєць, а ще три — 62, 56 та 66. Всі ці самиці мали яєць більше, ніж їхні яйцеводи могли б вигодувати ембріонів. В той час, як 5 ембріонів з самки, що експонувалась в AMNH, мали великий жовтковий міхур, 26 ембріонів з самки, впійманої біля узбережжя Мозамбіку, були близькі до народження, і мали лише шрам на животі в тому місці, де раніше був жовтковий міхур. Всі знайдені ембріони мали добре розвинуті травну систему та зуби. Таким чином, виглядає можливим, що додаткове харчування ембріонів відбувається за рахунок залишків надлишкових яєць. Окрім всього, відомо, що у деяких видів акул ембріони харчуються яйцями та іншими ембріонами, і врешті-решт народжується лише один великий хижак. Можливо, що така оофагія відбувається і у целакантів.
Подальші дослідження вищезазначених ненароджених ембріонів викрили надзвичайно широкі мембрани, що покривають жабри, і містять числені клітини, пристосовані до абсорбції внутрішньоутробного молока (гістотрофи), що секретується стінками яйцеводу. Цей тип передачі поживних речовин відомий і у деяких інших риб. Нарешті, каротиноїдні пігменти в жовтку також залучені до транспорту кисню.
Хоча для роз'яснення всіх питань в сфері розмноження потрібні додаткові дослідження, вже можна сказати, що целаканти — це риби з дуже розвиненою та складною репродуктивною системою. Це, одначе, не стало для вчених сюрпризом, з огляду на те, що юрський целакант Holophagus gulo був, вірогідно, живородящим, а целакант з Кам'яновугільного періоду Rhabdoderma exiguum, хоч і розмножувався овіпорозом, але мав яйця великого розміру та складної структури.
Згідно до непрямих відомостей, вагітність целакантів дуже довга (близько 13 місяців), самиці стають статево зрілими у віці більше ніж 20 років (як у деяких осетрових), і після досягнення статевої зрілості розмножуються один раз на кілька років. Досі невідомо, як відбувається внутрішнє запліднення, і де живуть молоді риби кілька років після народження. При зануреннях жодної молодої риби не спостерігали біля берегів або в печерах, і тільки дві були знайдені вільно плаваючими в товщі води.
http://uk.wikipedia.org/wiki/Целакант